Location: Home >> Detail
Crop Breed Genet Genom. 2025;7(3):e250009. https://doi.org/10.20900/cbgg20250009
 ,
Peter Dapprich 1 ,
Hendrik Bertram 1,2 ,
Jacqueline Hollensteiner 1 ,
Michaela Schmitz 1 ,
Armin O. Schmitt 2,3 ,
Mehmet Gültas 1,3,*
,
Peter Dapprich 1 ,
Hendrik Bertram 1,2 ,
Jacqueline Hollensteiner 1 ,
Michaela Schmitz 1 ,
Armin O. Schmitt 2,3 ,
Mehmet Gültas 1,3,*
1 Faculty of Agriculture, South Westphalia University of Applied Sciences, Lübecker Ring 2, Soest 59494, Germany
2 Breeding Informatics Group, Georg-August University, Margarethe von Wrangell-Weg 7, Göttingen 37075, Germany
3 Center for Integrated Breeding Research (CiBreed), Georg-August University, Albrecht-Thaer-Weg 3, Göttingen 37075, Germany
* Correspondence: Mehmet Gültas
Climate change amplifies drought stress, threatening global crop yields and highlighting the urgent need for effective soil amendment strategies. SAPs are increasingly proposed as soil amendments to improve drought resilience, although their agronomic performance across species and trait categories remains insufficiently characterized. We systematically evaluated three SAP types—one biopolymer-based and two fossil-based—across 14 morphological, physiological, and biochemical traits in two maize and two rapeseed genotypes under controlled drought conditions. PCA and a RF-based feature selection identified SFW, PH, and total PFW as key responsive traits. However, SAP applications did not significantly enhance biomass accumulation or antioxidant activity, with pronounced genotype-specific responses, particularly in rapeseed. Fossil-based SAPs consistently outperformed the biopolymer-based variant. Our results demonstrate that SAP efficacy is species- and genotype-dependent, challenging the assumption of its universal benefit under drought. This multi-trait, dual-species analysis underscores the need for genotype-specific SAP strategies and highlights the value of machine learning approaches for predicting treatment outcomes. Overall, our findings contribute to a more nuanced understanding of SAP–soil–plant interactions and support the targeted development of climate-resilient agricultural technologies.
SAP: superabsorbent polymer; ABG: agrobiogel; SWT: isonem soil water trap; TPC: total phenol content; PCA: principal component analysis; RF: random forest; SH: shoot height; RL: root length; RFW: root fresh weight; SFW: shoot fresh weight; RDW: root dry weight; SDW: shoot dry weight; PH: plant height; PFW: plant fresh weight; PDW: plant dry weight; LA: leaf area; RA: root area; SWC: shoot water content; RWC: root water content.
The rapid acceleration of global warming and climate change presents complex challenges to sustainable agriculture, crop protection, and global food security [1,2]. The consequences of these phenomena contain both abiotic and biotic stresses. Abiotic stresses, including environmental factors such as drought, temperature fluctuations, salinity, and heat [1–3], can decrease plant production by up to 70% [4]. Among these stressors, drought, characterized as a natural phenomenon resulting in insufficient water availability for human and ecosystem needs [5,6], significantly influences agriculture, leading to potential reductions in plant growth, plant health [7,8], and diminished yields [9]. Hence, global efforts to develop strategies for improving water use efficiency are intensifying [10].
One promising approach to coping with drought stress in agriculture involves utilizing innovative materials such as SAPs [11]. SAPs, characterized as three-dimensional hydrophilic polymers with a cross-linked network structure, exhibit the remarkable ability to absorb water hundreds to thousands of times their weight [12–14]. Great efforts have been made in the last years for the extensive usage of SAPs in agriculture, primarily as water and nutrient storage and retention materials, as well as additives that improve soil properties for various plant applications [14,15]. These polymers might function as a rhizosphere water reservoir, leading to drought stress resistance, modifying soil structure, and enhancing fertilizer efficiency in the agricultural sector [16–18]. SAPs can be classified into two main categories: natural or biopolymer-based and chemical/fossil-based. While fossil-based SAPs, so-called synthetic polymers that are sodium polyacrylate, suffer from drawbacks such as environmental pollution, natural SAPs are derived from polysaccharides and polypeptides, including cellulose, starch, agarose, chitosan, and their derivatives, which avoid such environmental troubles [19,20]. Despite several positive outcomes of applying this material to the soil, there are also notable disadvantages. Sepehri et al. [21] suggest that uncertainty exists regarding water accessibility for plants when SAPs are utilized. This may lead to competition between plants and SAPs for water absorption, potentially reducing the amount of water available for plants [22]. Moreover, studies by Zhang et al. [23] and Liu et al. [24] have indicated that SAPs can absorb various cations such as Na+, which may subsequently be taken up by plants through their roots, potentially affecting plant growth dynamics [22]. Furthermore, the water absorption capacity of SAPs is influenced by factors such as soil structure, temperature, pH, the soil wetting-drying cycle, and mineral elements in the water [11,25]. Therefore, it is essential to thoroughly investigate the interaction mechanisms to predict the conditions under which SAPs can enhance plant growth and drought resistance, as well as negative interference with plant growth by the accumulation of cations like Na+.
Most previous studies have focused on specific phenotypic traits, such as the effects of SAPs on physiological attributes, including chlorophyll content, PH, leaf number, stem diameter, and fresh and dry weights of shoots and roots in tomato [26]. Furthermore, yield and water use efficiency in rapeseed and wheat [27,28], and free proline content and total soluble sugars in rice leaves [29] have been investigated. Other studies have explored grain and straw yields in durum wheat [30] and winter wheat yield [31]. Additionally, germination rate and plant growth in wheat and cucumber [29], SH, RL, fresh and dry weights in barley [32], stem diameter, tuber yield, LA index, and percentage of dry matter in potato [33], as well as stem diameter, PH, and shoot and root biomass in spinach [34] have been studied.
Despite the extensive literature on applying SAPs in investigating specific plant traits, there is still a need for a comprehensive study. On the one hand, such a study should evaluate the overall impact of different SAP types on the three key trait groups—morphological, physiological, and biochemical. On the other hand, it should systematically explore genotype-specific responses across different cultivars of major crops such as maize and rapeseed, thereby enabling more targeted and efficient applications of SAPs in crop improvement strategies. To address this gap, our study adopts a novel dual-species approach using maize and rapeseed, enabling a comparative analysis of SAP efficacy across different cultivars. By integrating machine learning techniques, we aim to identify the most responsive traits affected by SAPs, offering a robust, data-driven understanding of genotype- and SAP-type-specific interactions. Based on these insights, our extensive and systematic analysis could enhance trait-based selection for breeding programs and contribute to developing drought-resilient crops. To this end, we conducted a greenhouse experiment to collect the corresponding phenotypical data of 14 traits, encompassing all three characteristic groups of rapeseed and maize plants, using three different SAPs (one biopolymer-based and two chemical/fossil-based). Following the experiment, we systematically evaluated trait responses by applying two complementary machine learning approaches—PCA and RF feature selection. PCA, as an unsupervised method, identified major sources of variation and trait co-variation across samples, while Boruta-RF, a supervised method, ranked traits by their importance in distinguishing between SAP treatments. This integrated approach enabled a comprehensive assessment of both general variability and treatment-specific effects. Interestingly, both methods consistently identified SFW, PH, and PFW as key indicators of SAP performance across species and cultivars, while also revealing species-specific responses—root and SDW in maize and LA in rapeseed—underscoring the differential trait responsiveness to SAPs. Significant differences across rapeseed cultivars were observed for most traits under SAP treatments, a pattern not consistently evident in maize. Among the SAPs, the biopolymer-based ABG generally exhibited lower performance relative to one or both fossil-based treatments, except for the traits SWC in rapeseed and RDW in maize. On top of that, our findings revealed that SAPs do not exhibit universal positive effects, with significant variation observed across treatments and control groups. The differential impacts of SAPs on oxidative stress responses further emphasized their conditional effectiveness under drought stress.
The investigation was executed in the glass greenhouse under a 16/8 h light/dark cycle at South Westphalia University of Applied Sciences, Soest, Germany (51°34′32.09″ N, 8°06′22.28″ E), in the summer and winter of 2023. The experimental design employed a completely randomized design incorporating two variables:
1.
In addition to investigating the impact of drought stress and the effects of SAPs in the experimental setting, two control groups (soil without SAPs) were incorporated. The first control group received regular irrigation, referred to as the control normal (CN). In the second group, water-stress conditions were induced to match the SAP treatment’s water regime, referred to as the control stress (CS).
2.
Regarding the absorption capacity (the amount of water, in mL, that one gram of SAPs can absorb) required to induce the same stress level in plants, it was necessary to verify the theoretical absorption capacity stated by the SAP producer in the product details. To do so, five repeated tests were conducted (Table 1). The results showed that the absorption capacity of MERCK SAP was almost the same as the theoretical capacity claimed by the company when using distilled water. In contrast, ABG demonstrated a minimum absorption of 10 times the amount of water claimed by the company. For SWT, the absorption capacity was 250 mL/g, which was 58% less than the theoretical absorption capacity. Since distilled water is generally not used in field and greenhouse experiments, we also tested the absorption capacity using tap water (commonly used in these experiments). The results showed a decrease in the absorption capacity of both fossil-based SAPs, with a 49% reduction for MERCK and a 72% reduction for SWT compared to the theoretical capacity. This trend was different for ABG, where the absorption capacity increased compared to distilled water and fell within the range of the theoretical absorption capacity. Given that fertilizers are typically used during experiments, it was important to assess the absorption capacity when fertilizers were applied. The absorption capacity decreased by more than 62% for MERCK and 80% for SWT when water was combined with fertilizer (NPK). For the natural-based SAP, we reached the maximum absorption capacity when using fertilizer. These results indicate that SAPs interact with water’s mineral content and the fertilizer’s cations and anions. The amount of SAPs used in this experiment was based on the recommended dosage, taking into account the results from this analysis.
 Table 1. Water absorption capacity of SAPs with varying water sources and fertilizers.
Table 1. Water absorption capacity of SAPs with varying water sources and fertilizers.
To ensure uniform stress levels across all plants, soil moisture was continuously monitored using the TEK-255G Indoor Outdoor Soil Moisture Meter, Tekcoplus Ltd, China. The monitoring continued until approximately 50% of the plants in the dry-level irrigation treatment (moisture level 3) exhibited relative wilting. Based on pre-tests, which accounted for soil texture, average greenhouse temperature, and pot size, a standardized 200 cc of water was administered to each pot for the rapeseed experiment. In contrast, the control group received regular irrigation to prevent drought stress. In the maize experiment, 500 cc of water was provided for each pot. To systematically study the effects of SAPs on both plant species, 14 distinct traits were measured and categorized into three groups (Figure 1). The first group consists of morphological characteristics, including SH, RL, RFW, SFW, RDW, SDW, PH, PFW, PDW, LA, and RA. The second group comprises physiological traits, namely SWC and RWC, along with the third group as biochemical trait, which is TPC. Measurements were taken using a ruler to measure traits related to length and height (measured in centimeters). Weights (measured in grams) were determined using a scale (Sartorius TE214S, Germany). The fresh organs were then dried in an oven at 70 °C for 72 h to measure dry weights [35–37]. SWC and RWC were calculated using the formula 1 [38]:
Where WC, in percentage, refers to water content, FW denotes fresh weight, and DW represents the dry weight of the shoot or root part.
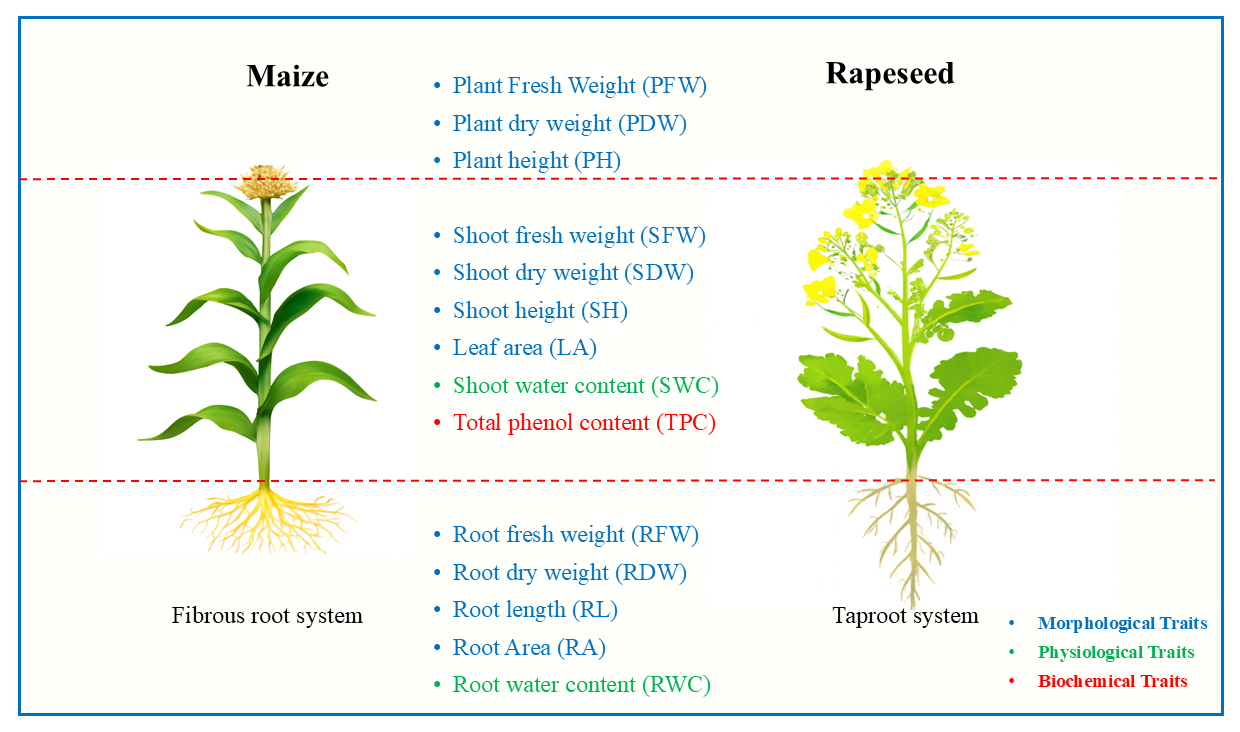 Figure 1. Schematic representation of the plant species and their 14 traits, categorized by phenotype groups. Traits are measured from various plant tissues, with color coding indicating their respective groups.
Figure 1. Schematic representation of the plant species and their 14 traits, categorized by phenotype groups. Traits are measured from various plant tissues, with color coding indicating their respective groups.
A total of 150 pots were prepared for each plant species (maize and rapeseed), providing fifteen replicates for every treatment level, including the two control conditions (CN and CS). For the assessment of morphological and physiological traits, we analyzed 120 pots per species, corresponding to twelve independent replicates of each variety under every treatment and control condition. For the biochemical determination of antioxidant capacity, we further selected 30 pots per species, representing three replicates for each treatment-variety combination, controls included. This tiered sampling scheme ensured adequate statistical power for the overall experiment while keeping the workload manageable for the labor-intensive phenotypic and biochemical analyses. A standardized soil mixture was prepared to ensure optimal plant growth conditions. The soil composition consisted of 30% sandy soil mixed with 70% peat moss. The sandy soil was sterilized in an autoclave for 90 min at 120 °C. The pH of peat moss was 5.8–6.8, and the soil consisted of 340 mg/L Nitrogen (N), 260 mg/L phosphate (P2O5), 330 mg/L (K2O, Potassium, and 100 mg/L magnesium (Mg). Each pot (12 cm × 12 cm × 12 cm) was filled with 1.5 kg of the soil blend, with one seed sown per pot for rapeseed. For maize, each pot (20 cm × 18 cm × 20 cm) was filled with 5 kg of the soil blend.
Antioxidant Extraction: TPC DeterminationOne gram of gallic acid was dissolved in 100 mL of methanol, resulting in a 1 % gallic acid solution (10 mg/mL) designated as the standard solution. To construct a standard gallic acid curve, dilutions (0.1, 0.2, 0.4, 0.6, 0.8, and 1 mg/mL) were prepared in methanol from the first standard solution (Solution 1). Each dilution (0.1 mL) was mixed successively with 0.5 mL of water and 0.1 mL of Folin-Ciocalteu reagent, followed by a 6 min incubation period. Subsequently, 1 mL of sodium hydroxide 1% and 0.5 mL of distilled water were introduced to the reaction mixture. The absorbance was recorded spectrometrically after 90 min at 760 nm (UV-1800 spectrophotometer, Shimadzu, Duisburg, Germany) [39]. Utilizing the standard gallic acid curve and regression equation, the total phenolic content of the extracts was calculated based on y = 0.0352 + 3.2098x (𝑅2 = 0.9976), where y is the absorbance and x is the concentration of dilutions, as shown in Figure 2.
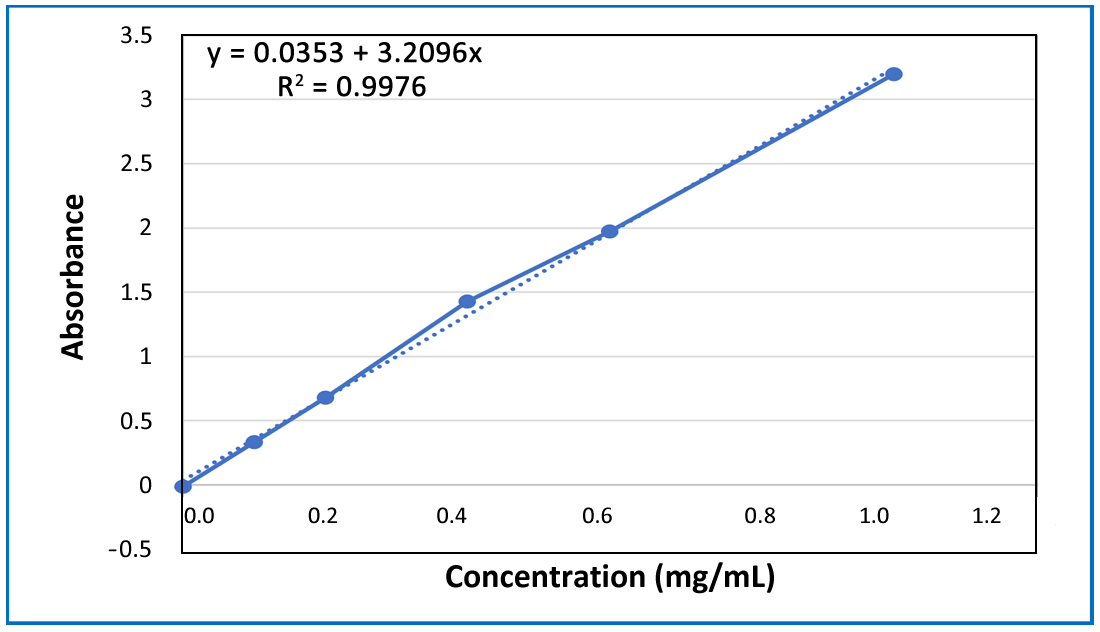 Figure 2. Standard curve of gallic acid: Calibration curve to measure the TPC concentration in plant samples.
Figure 2. Standard curve of gallic acid: Calibration curve to measure the TPC concentration in plant samples.
The TPC is extracted from plant samples by the following procedure: 200 mg of true leaves were immersed in liquid nitrogen for 3 min, followed by mechanical milling for 10 min at 25,000 rpm (Retsch Mixer Mill MM200, Haan, Germany). The resulting milled sample was then transferred into a centrifuge tube, and methanol (3 × 1 mL) was added. Subsequently, centrifugation was conducted at 5000 rpm and 4 °C for 5 min (Rotina 380R, Hettich, Kirchlengern, Germany) after each methanol addition. A mixture was prepared by combining 0.5 mL of deionized water, 0.5 mL of Folin-Ciocalteu, and 0.5 mL of the sample for 30 s. Then, 5 mL of 1% sodium hydroxide was added, and the mixture was vortexed. The resulting solution was then incubated precisely for 30 min, after which its absorbance was measured at 𝜆 = 760 nm using a UV spectrophotometric device (UV-1800 spectrophotometer, Shimadzu, Duisburg, Germany) [39].
Quantification and Statistical AnalysesData are primarily depicted as box plots, with mean values shown by crosses and individual data points overlaid. The analyses were conducted using R software (version 4.2.3 [40]). To assess significant differences between groups (both treatment groups and cultivar groups), we applied either a one-way ANOVA with Tukey’s test as post hoc test or the Kruskal-Wallis test followed by Dunn’s test as post hoc test, depending on the normality of the data distribution which was assessed using the Shapiro-Wilk test [41]. The significance threshold for all analyses was established at α = 0.05.
Furthermore, two specific traits, LA and RA, were quantified using image analysis. Images were captured using a Nikon camera (Nikon D5100 + AF-S DX Nikkor, Bangkok, Thailand), and measurements were obtained using the ImageJ software (Version 1.54d) [42,43]. Additionally, a custom Python script and the LeafArea package (Version 0.1.8) [44] in R were utilized to further analyze and process the obtained data. Phenotypic correlations were calculated based on the Pearson correlation coefficient (r) between each pair of quantitative traits to assess their relationships in both plant species. In Figure 3, a sample of the image analysis results for both plants is shown. This figure highlights the differences in root architecture between maize, which has a fibrous root system with several main primary roots, and rapeseed, which features a taproot system with one main primary root and several lateral roots. Additionally, the figure illustrates the impact of the treatments, showing how the SAP application affects LA, RA, and, consequently, the fresh and dry biomass weight.
 Figure 3. RA and LA for both plant species. Binary images of RA and LA for (A) maize and (B) rapeseed, which were used to calculate RA and LA. In (A) and (B), the upper panels display the root architecture of each species, while the lower panels show the corresponding LA. These images illustrate the variations in root and leaf morphology between maize and rapeseed under different treatments.
Figure 3. RA and LA for both plant species. Binary images of RA and LA for (A) maize and (B) rapeseed, which were used to calculate RA and LA. In (A) and (B), the upper panels display the root architecture of each species, while the lower panels show the corresponding LA. These images illustrate the variations in root and leaf morphology between maize and rapeseed under different treatments.
The systematic assessment of these traits based on their potential for SAP application is crucial for guiding further investigations, especially when selecting target phenotypes (traits) for breeding programs. To this end, we applied two machine learning methods: PCA, (R packages: factoextra [45] and gridExtra [46]), the RF, along with the Boruta algorithm [47] to investigate the importance of traits influenced by the SAP application. As an unsupervised learning method, PCA was performed to reduce the dimensionality of the complex datasets while preserving variance, thus enabling the identification of key traits by assessing their contributions to the principal components. This method is used to identify key variables that contribute to the observed phenotypes [48].
The RF algorithm, a supervised learning method, was employed to estimate the relative importance of each trait in predicting response variables, which in this case were the SAP treatments and two control groups. To enhance feature selection, we applied the Boruta algorithm [47], a specialized wrapper tailored for feature selection using the RF framework. The algorithm works by augmenting the dataset with additional random attributes, called shadow attributes, created by shuffling the original trait values. This process introduces controlled randomness, reducing biases from false signals caused by random fluctuations or spurious correlations in the data. An RF classifier is then employed on the extended dataset to evaluate the importance of each trait. Traits with importance scores significantly lower than those of the shadow attributes are iteratively removed. By repeatedly generating shadow attributes and applying the RF classifier, the Boruta algorithm assigns important scores to all traits. Ultimately, it ranks the traits and classifies them as confirmed, rejected, or tentative based on their importance [49,50].
With the growing challenges posed by climate change, developing innovative agricultural solutions, such as SAPs, has become increasingly critical to enhance crop resilience. Despite their potential, the impact of different SAPs on key plant traits and their specific role in plant development remain poorly understood. In this study, we systematically evaluated 14 traits in maize and rapeseed to investigate the impact of SAP application on both crops. Through comparative analyses, our findings provide novel insights into the potential of these traits to mitigate climate-related stresses and improve crop productivity.
Assignment of the Importance of Individual TraitsUnderstanding which plant traits are most influenced by SAP application is essential for guiding future breeding programs and enhancing crop performance. While several studies [51–53] have demonstrated the potential of SAPs to improve plant yield and trait development, a more holistic approach is required to rank these traits and clarify their roles in SAP-driven improvements. To address this, we first applied the RF approach, utilizing the Boruta algorithm, to the datasets from maize and rapeseed. This analysis identified five traits—SFW, PH, SDW, PFW, and PDW—in maize and five traits—PFW, SFW, PH, LA, and RL—in rapeseed as important for predicting the response variables (treatments: SAPs and both control groups) (Figure 4A,B).
Moreover, PCA analysis further highlighted the most important traits in both datasets based on their contributions to the principal components. For maize, the traits PDW, SDW, PFW, SFW, PH, RL, RDW, and LA were identified as key contributors (Figure 4C). Similarly, for rapeseed, the traits PDW, PFW, SFW, SDW, RFW, LA, RDW, RA, and PH were deemed most important (Figure 4D). Interestingly, among the important traits, SFW, PH, PFW, PDW, and SDW in maize and PFW, SFW, PH, and LA in rapeseed were identified as common traits by both methods. This highlights their high relevance within the datasets, representing key traits for SAP applications in each plant species.
A closer look at these traits assigned as important by RF or PCA for both maize and rapeseed suggests that, regardless of the machine learning approach used, the traits PH, PFW, and SFW consistently play a crucial role in characterizing the effects of the SAP application. This indicates that these traits exhibit reliable and consistent behavior across different plant species under drought stress conditions when SAPs are applied. In this regard, the importance of PFW and SFW could be explained by their biological relevance in determining overall plant biomass and their strong association, as shown in the section “Analysis of Associations between Quantitative Traits”. This finding aligns with our expectations, reinforcing the idea that changes in one trait often correspond to changes in the other. Additionally, the traits commonly identified by RF and PCA revealed species-specific patterns; RDW and SDW were exclusive to maize, whereas LA was unique to rapeseed. These findings indicate that SAPs affect some traits similarly across species while also eliciting distinct responses depending on the plant type.
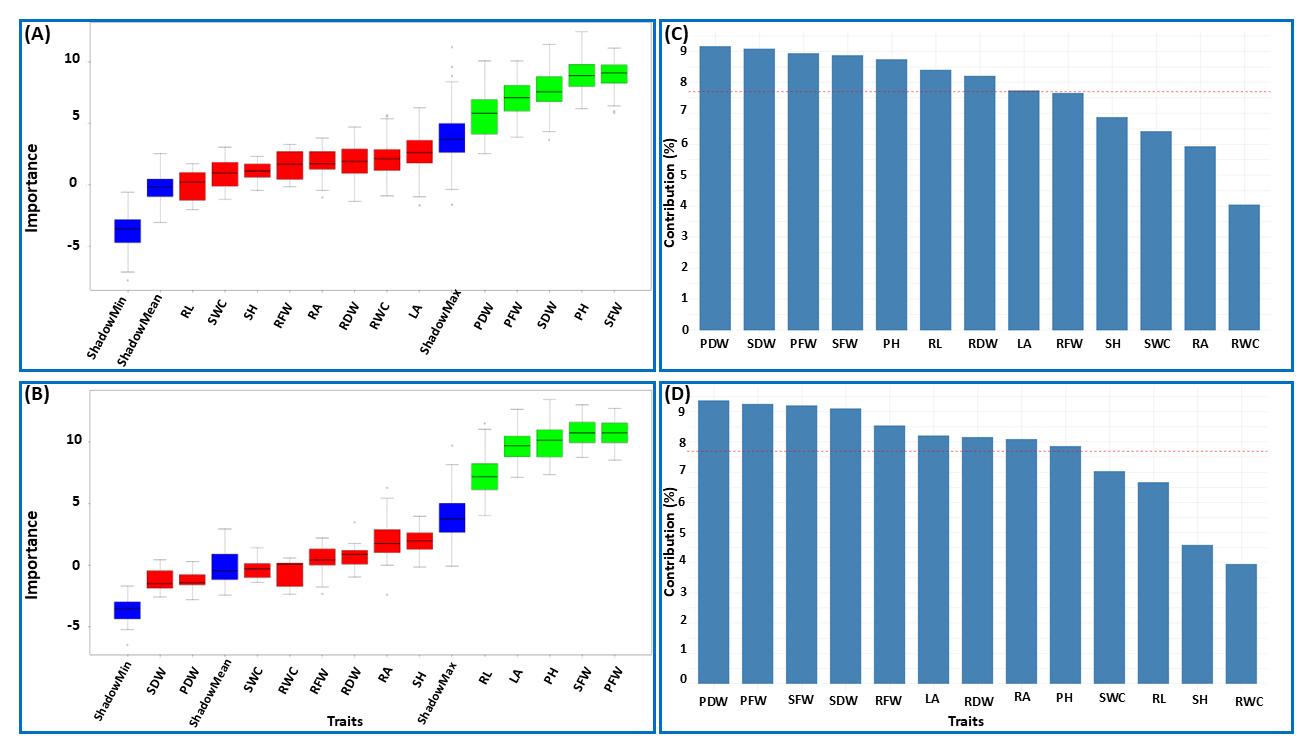 Figure 4. Importance of individual traits. Boruta analysis of trait importance for maize (A) and rapeseed (B), where green and red boxes indicate confirmed and rejected trait importance, respectively. The blue boxes refer to shadow features (max, min, and mean) which are generated by permuting the original values, providing a baseline for identifying important traits. PCA-based trait importance for maize (C) and rapeseed (D), with traits above the red dashed line (mean importance) regarded as important.
Figure 4. Importance of individual traits. Boruta analysis of trait importance for maize (A) and rapeseed (B), where green and red boxes indicate confirmed and rejected trait importance, respectively. The blue boxes refer to shadow features (max, min, and mean) which are generated by permuting the original values, providing a baseline for identifying important traits. PCA-based trait importance for maize (C) and rapeseed (D), with traits above the red dashed line (mean importance) regarded as important.
The application of machine learning methods revealed three traits—SFW, PFW, and PH—as important in both maize and rapeseed. To assess the impact of SAP application and drought stress, we focus on a detailed analysis of these traits. Due to the strong correlation between SFW and PFW (r = 1.00) in both plant species (See Section Analysis of Associations between Quantitative Traits), the analysis primarily concentrates on SFW and PH. Further results for the remaining traits are provided in Supplementary Material Figures.
SFWThe water deficit in plants typically appears as a decrease in shoot weight, especially in fresh weight, which in turn affects the overall dynamics of shoot growth. As a result, changes in SFW serve as a reliable indicator of a plant’s response to drought stress. The results indicate distinct responses to treatments between maize and rapeseed regarding SFW. In maize (Figure 5A), a significant reduction in SFW was observed in all SAP and CS treatments compared to the CN, suggesting that the application of SAPs did not enhance shoot biomass under the tested conditions. For rapeseed, as shown in Figure 5B, the only significant difference was between MERCK and CN, where the MERCK treatment resulted in lower SFW compared to the control. Other SAP treatments did not show significant differences from the control, indicating that rapeseed was generally less responsive to SAP applications in terms of SFW. When comparing cultivars across species (Figure 5C), no significant differences were found between maize cultivars in any treatment, indicating that SAPs had a uniform effect on maize regardless of variety. In contrast, significant differences were observed between rapeseed cultivars for all treatments, suggesting that genetic background played a crucial role in determining SFW in rapeseed. These findings highlight species-specific and cultivar-specific responses to SAP applications.
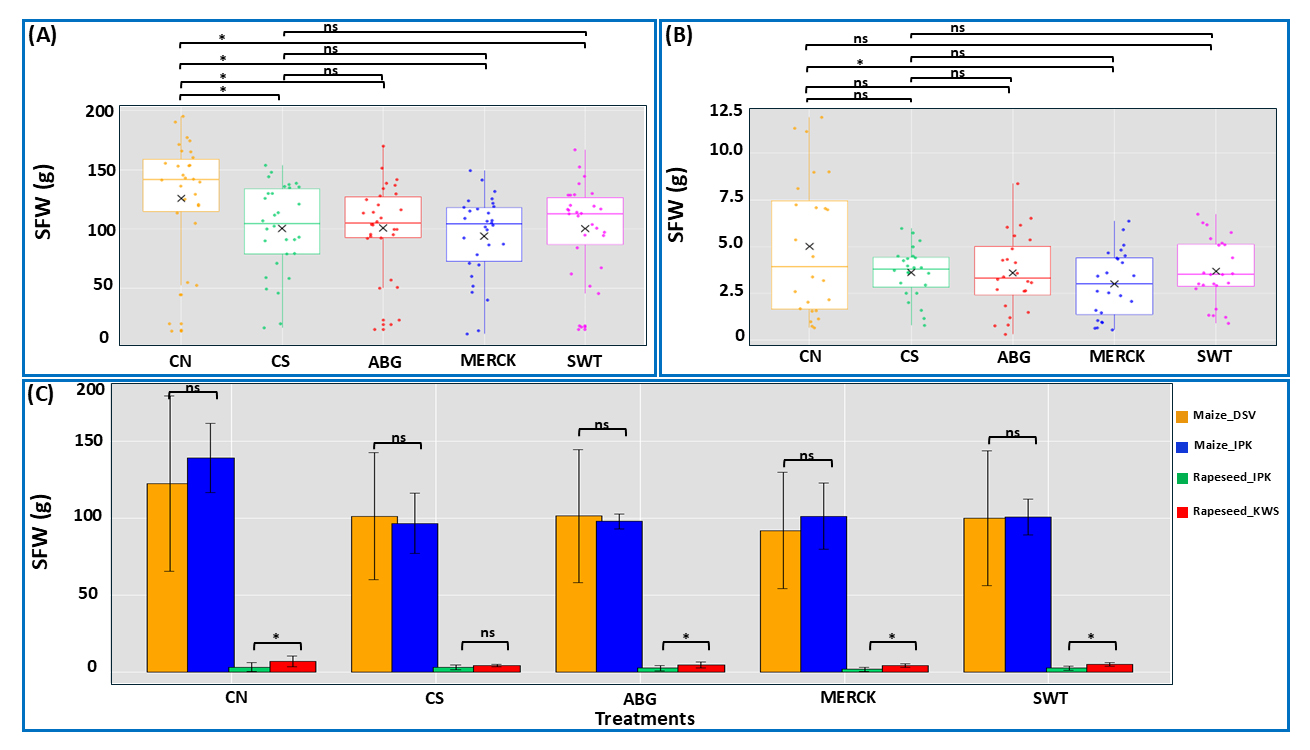 Figure 5. SFW analysis. Box plots depict SFW in maize (A), rapeseed (B), and cultivar comparisons across both plant species (C). Panels A and B depict treatment effects (treatments: SAPs and both control groups) independent of cultivar differences, while panel C illustrates the influence of cultivar on treatment outcomes Statistical analysis was performed using the Kruskal-Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment. In panel C, the colors represent the following genotypes: orange color stands for the maize genotype provided by DSV company, blue color stands for the maize genotype from IPK, green color stands for rapeseed genotype provided by IPK, and red color stands for the rapeseed genotype from KWS company.
Figure 5. SFW analysis. Box plots depict SFW in maize (A), rapeseed (B), and cultivar comparisons across both plant species (C). Panels A and B depict treatment effects (treatments: SAPs and both control groups) independent of cultivar differences, while panel C illustrates the influence of cultivar on treatment outcomes Statistical analysis was performed using the Kruskal-Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment. In panel C, the colors represent the following genotypes: orange color stands for the maize genotype provided by DSV company, blue color stands for the maize genotype from IPK, green color stands for rapeseed genotype provided by IPK, and red color stands for the rapeseed genotype from KWS company.
In conditions of limited water availability, plants often exhibit diminished vertical growth, reducing overall PH. In maize (Figure 6A), PH was significantly lower in all stress situations, including SAP treatments and CS group (ABG, MERCK, SWT, and CS) compared to the well-watered control (CN), indicating that drought stress, despite of SAP application, negatively affected growth. However, SAPs did not alleviate this effect, as no statistically significant differences were observed among the stress-treated groups. In rapeseed (Figure 6B), a significant reduction in PH was observed only in the ABG treatment compared to both control groups (CN and CS), whereas MERCK and SWT showed no significant differences, suggesting minimal impact on PH. Regarding cultivar comparisons (Figure 6C), a significant difference between maize cultivars was detected only under CN, while both cultivars responded similarly under stress conditions. In rapeseed, cultivar differences were significant in all treatments except ABG, highlighting a more pronounced cultivar effect under SAP treatments. Overall, maize PH was highly sensitive to drought stress, with all stress treatments leading to a significant reduction in height compared to normal conditions, and SAPs failing to mitigate this decline. In rapeseed, ABG negatively affected PH, whereas other SAP treatments had no significant impact. Cultivar effects were more pronounced in rapeseed, whereas in maize, differences were only evident under well-watered conditions.
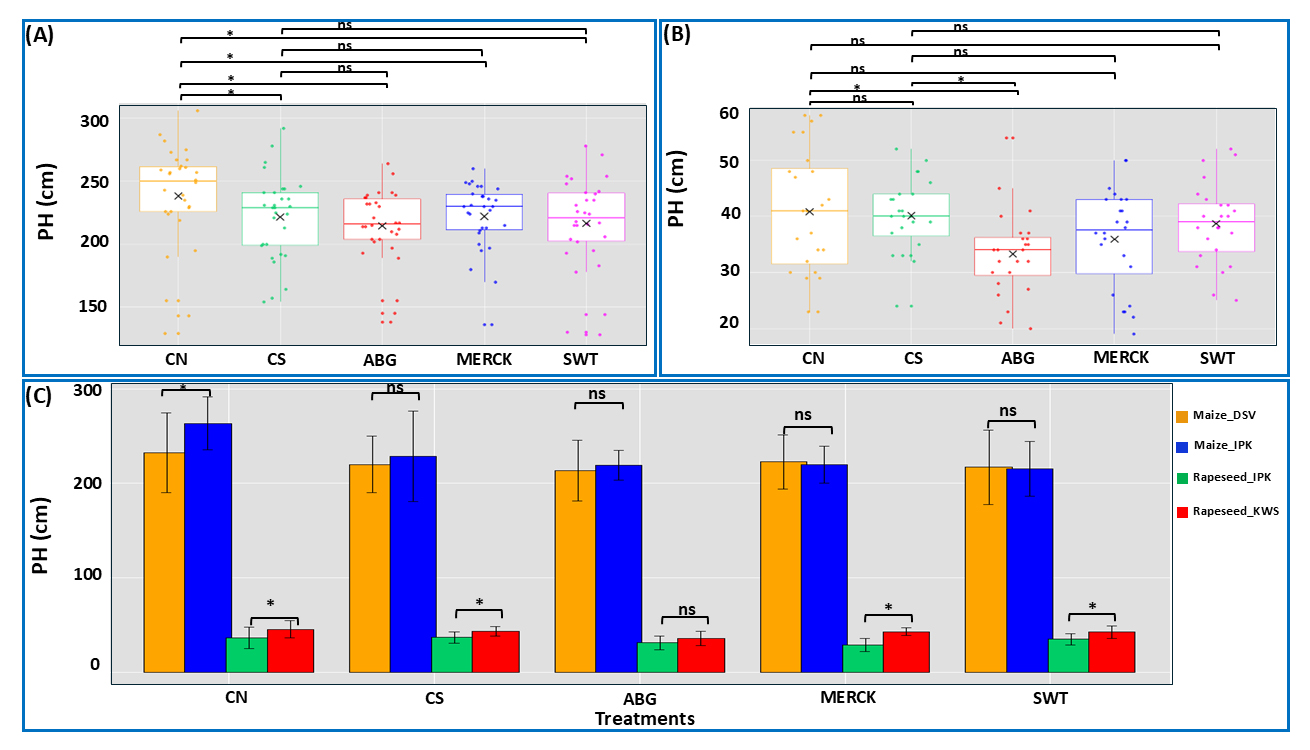 Figure 6. PH analysis. Box plots display PH in maize (A), rapeseed (B), and cultivar comparisons across both plant species (C). Panels A and B depict treatment effects (treatments: SAPs and both control groups) independent of cultivar differences, while panel C illustrates the influence of cultivar on treatment outcomes. For the maize plots, statistical analysis was performed using the Kruskal–Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. Statistical analysis for rapeseed was conducted using one-way ANOVA, followed by Tukey’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment. In panel C, the colors represent the following genotypes: orange color stands for the maize genotype provided by DSV company, blue color stands for the maize genotype from IPK, green color stands for rapeseed genotype provided by IPK, and red color stands for the rapeseed genotype from KWS company.
Figure 6. PH analysis. Box plots display PH in maize (A), rapeseed (B), and cultivar comparisons across both plant species (C). Panels A and B depict treatment effects (treatments: SAPs and both control groups) independent of cultivar differences, while panel C illustrates the influence of cultivar on treatment outcomes. For the maize plots, statistical analysis was performed using the Kruskal–Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. Statistical analysis for rapeseed was conducted using one-way ANOVA, followed by Tukey’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment. In panel C, the colors represent the following genotypes: orange color stands for the maize genotype provided by DSV company, blue color stands for the maize genotype from IPK, green color stands for rapeseed genotype provided by IPK, and red color stands for the rapeseed genotype from KWS company.
TPC was determined using the calibration curve (See Antioxidant extraction: TPC determination) and expressed in mg/mL gallic acid equivalent. In maize, no significant differences in TPC were observed between SAP treatments (ABG, MERCK, SWT) and CN (Figure 7A). While the SAP treatments, especially those with MERCK, showed a slight decrease in TPC under drought stress conditions compared to the two control groups, these differences were not statistically significant. However, in rapeseed, a distinct response was observed (Figure 7B), where TPC levels were significantly higher in the ABG compared to CN, suggesting that the natural-based SAP (ABG) promoted phenolic accumulation. In contrast, MERCK and SWT treatments did not significantly differ from CN, indicating that these SAPs had no substantial effect on TPC synthesis. Plants are required to adjust to fluctuations caused by abiotic factors. The accumulation of phenolic compounds in plant tissues is believed to be an adaptive mechanism in response to these adverse environmental conditions. The increase in TPC under stress conditions reinforces the well-documented role of drought stress in stimulating secondary metabolite production, a common physiological response linked to enhanced antioxidative defense mechanisms in plants [54]. Conversely, a decrease in TPC in the treatments indicates that the plants are experiencing less stress.
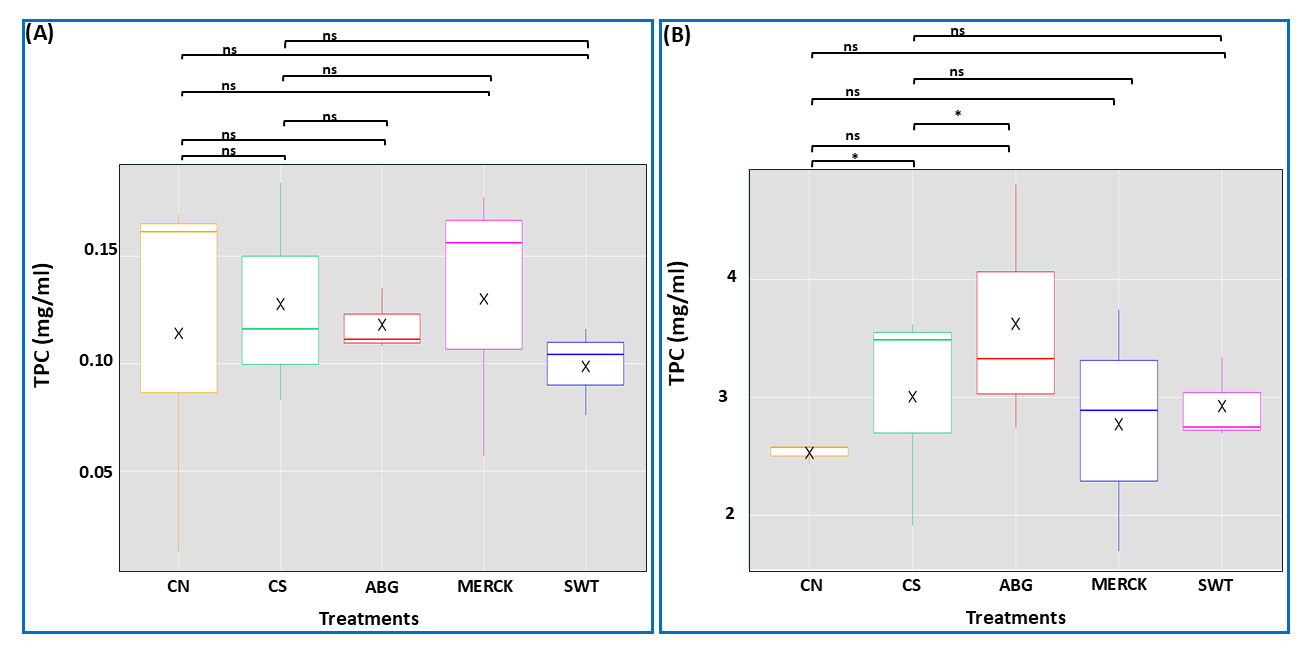 Figure 7. TPC analysis. The effects of treatments and control groups on TPC in maize (A), and rapeseed (B). For maize, statistical analysis was performed using one-way ANOVA, followed by Tukey’s test for post hoc pairwise comparisons. Statistical analysis for rapeseed was conducted using the Kruskal-Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment.
Figure 7. TPC analysis. The effects of treatments and control groups on TPC in maize (A), and rapeseed (B). For maize, statistical analysis was performed using one-way ANOVA, followed by Tukey’s test for post hoc pairwise comparisons. Statistical analysis for rapeseed was conducted using the Kruskal-Wallis test, followed by Dunn’s test for post hoc pairwise comparisons. The symbol ‘ns’ denotes no significant difference between comparison pairs, and ‘*’ indicates significant differences at α = 0.05. The ‘×’ symbol represents the mean value for each treatment.
We analyzed the data using the Pearson correlation coefficient (r) to assess the strength of relationships among the quantitative traits of both plants, combining data from both control groups and SAP treatments. As illustrated in Figure 8, the results revealed a consistent positive correlation among most of the traits examined. However, physiological traits RWC and SWC showed negative correlation coefficients with the other traits, distinguishing them from the overall trend. Notably, the correlation values indicate particularly strong associations between specific trait pairs. For instance, a strong positive correlation was observed between RA and PFW, with coefficients of 0.75 in maize and 0.82 in rapeseed. Similarly, the relationship between RDW and SDW exhibited high correlation coefficients of 0.91 in maize and 0.81 in rapeseed, underlining the tight association between these traits. Moreover, LA exhibited a strong positive correlation with RA, with coefficients of 0.82 in rapeseed and 0.73 in maize. Similarly, LA was strongly correlated with RFW, with correlation coefficients of 0.82 in maize and 0.71 in rapeseed, further highlighting the interdependence among key phenotypic traits. Interestingly, particular trait correlations, such as those between PFW and LA (r = 0.91), as well as SFW and PH (r = 0.80), were identical across both plant species. This consistency indicates that the SAP application does not alter the inherent relationships between these traits.
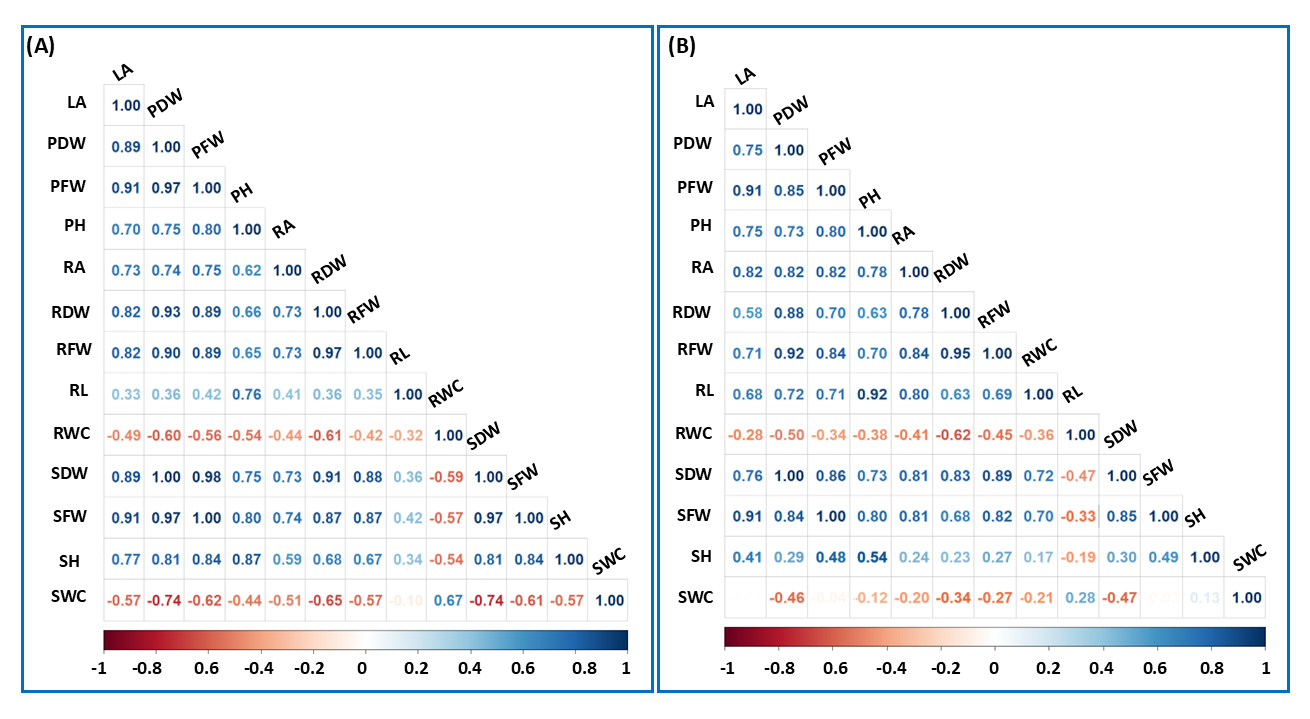 Figure 8. Correlation heatmap across traits. Correlation heatmap for maize (A) and rapeseed (B), where the intensity of blue and red indicates the strength of positive and negative correlations, respectively.
Figure 8. Correlation heatmap across traits. Correlation heatmap for maize (A) and rapeseed (B), where the intensity of blue and red indicates the strength of positive and negative correlations, respectively.
Plants deploy diverse physiological and biochemical mechanisms to cope with environmental stressors, including biotic and abiotic challenges [9]. Among these, drought stress has been extensively studied due to its detrimental impact on plant growth, morphology, and biochemical processes, ultimately resulting in substantial yield losses [55–60]. In this context, SAPs have emerged as promising tools for alleviating drought-induced stress. Several studies have discussed that SAPs could partially mitigate the adverse effects of water scarcity by improving water availability and enhancing plant stress tolerance. Regarding the effects of SAPs, previous studies can be roughly categorized into two main groups: (i) those reporting beneficial impacts on plant performance under drought stress, and (ii) those indicating neutral or even negative effects, highlighting the context-dependent nature of SAP efficacy. Among the documented benefits, SAPs have been shown to improve soil structure [61], enhance water retention [62,63], and enable slow-release behavior [64–66] when applied to soil. These improvements have translated into enhanced physiological parameters in various crops, such as increased germination rates, improved plant growth [17,67,68], root system development [6,69], larger LA [70], higher oil content [29], and greater yield and biomass in several crops and vegetables [71–74]. Conversely, other studies have identified notable limitations associated with SAP applications. These include competition for water between SAPs and plant roots [21], the release of sodium ions, and disruptive cation interactions in the soil [23,25]. However, these studies from both groups were typically restricted to a single plant species, examined a narrow range of phenotypic traits, and frequently involved only one SAP type, limiting their generalizability across crop types and environmental conditions.
To broaden the understanding of SAP performance beyond these limitations, we investigated two physiologically distinct crop species—maize (a monocot with a fibrous root system) and rapeseed (a dicot with a taproot system)—which differ in their root architecture, water uptake dynamics, and stress adaptation mechanisms. We employed a comprehensive phenotyping approach encompassing a wide range of morphological, physiological, and biochemical traits measured at the whole-plant level and the level of individual plant parts. This design allowed us to capture both general and species-specific responses to SAP treatments. Additionally, by including two genetically diverse cultivars per species, we revealed genotype-dependent variation in SAP efficacy, underlining the importance of intraspecific diversity in shaping treatment outcomes. Finally, the parallel evaluation of natural (biopolymer-based) and synthetic (fossil-based) SAPs under identical controlled drought conditions enabled a direct comparison of their functional effects, providing novel insights into their respective benefits and limitations. This integrative framework contributes to a more nuanced and generalizable understanding of SAP-plant interactions, with potential implications for future crop management and breeding strategies under water-limited conditions.
Our findings agree with previous reports by Akram et al. [75], who reported a decline in growth-related traits, including shoot and root fresh and dry weight, as well as total phenolic content, under drought conditions. Similarly, reductions in LA observed in our study align with prior investigations [76–79]. Moreover, Rafique et al. [80] documented a decline in plant fresh and dry biomass, with lower values under drought stress compared to control conditions, further confirming the impact of water deficit on plant growth. In our study, the observed decrease in biomass, reflected by significant reductions in SDW, RDW, SFW, and RFW, and consequently in PDW and PFW, is consistent with their findings and highlights the suppressive effect of drought on plant biomass accumulation. Notably, a significant reduction in maize biomass across all SAP treatments, as well as in the MERCK treatment for rapeseed, indicates that the SAPs, including the fossil-based MERCK, were insufficient to protect plants against drought-induced biomass loss under the tested conditions. The lack of significant improvement in most assessed traits following SAP application, coupled with an increase in antioxidant activity, reinforces the conclusion that SAPs did not effectively mitigate drought-induced oxidative stress. Overall, these results suggest that the detrimental effects of drought stress outweighed any potential positive effects of SAP treatments in both species.
Our findings regarding these traits concur with previous studies, which reported reductions in RL, SH, and PH were evident under drought stress, aligning with previous observations in rapeseed [81–84] and maize [85–87]. Furthermore, Rao et al. [88] demonstrated that barley cultivars exhibited significant differences in phenolic content and antioxidant production, a trend that was also observed in this study, particularly in rapeseed. Water deficit conditions have been shown to impair photosynthetic efficiency significantly [89,90]. Under such conditions, rapeseed has been reported to reduce leaf biomass by up to 34% [91] and decrease LA [92,93] as an adaptive strategy to minimize water loss via stomatal regulation. This effect was similarly observed in the present study. Drought-induced modifications in root structure and growth patterns have also been reported [89], and our results suggest that, despite SAP application, plants exhibited a stress response comparable to those observed in previous studies without SAP treatments. This suggests that SAPs had a limited effect on improving drought tolerance for the evaluated morphological traits under the tested conditions.
Plants exposed to abiotic stress, such as drought, experience oxidative stress primarily due to the excessive accumulation of reactive oxygen species (ROS), leading to lipid peroxidation, protein oxidation, and disruptions in cellular redox homeostasis [89]. To mitigate ROS-induced cellular damage, plants activate antioxidative defense mechanisms in which phenolic compounds and flavonoids serve as crucial non-enzymatic antioxidants, directly scavenging ROS and maintaining redox balance [94]. Consistent with these protective responses, Ayyaz et al. [95] reported enhanced antioxidant production in drought-stressed rapeseed, and Zhang et al. [96] observed elevated TPC in maize kernels under drought conditions. In accordance with these findings, our study revealed an increase in TPC under drought conditions in rapeseed. However, despite SAP applications, no significant reduction in TPC levels was observed, suggesting that SAPs did not sufficiently alleviate oxidative stress or suppress ROS accumulation in rapeseed. The persistent elevation of TPC under ABG treatment likely reflects ongoing activation of phenolic-based antioxidant defenses, indicating that the drought-induced oxidative stress was not fully mitigated. This may be attributed to species-specific responses, as rapeseed, a dicot with a deep taproot system, may sustain stronger oxidative signaling and phenolic metabolism even under marginal improvements in water availability. Moreover, limited SAP effectiveness could be associated with differences in polymer composition, ion exchange behavior, and restricted water release dynamics in the greenhouse environment. Interestingly, in maize, a decrease in TPC was observed under the MERCK treatment compared to the controls. This suggests that the MERCK SAP, possibly through improved osmotic regulation and water availability, was more effective in reducing ROS generation and oxidative stress in maize. Given the fibrous root system and differing stress adaptation mechanisms of maize, the species may have benefited more efficiently from SAP-mediated water retention. Furthermore, fossil-based SAPs like MERCK may interact differently with soil ions, supporting a better hydration status and thereby suppressing ROS-triggered phenolic accumulation. This finding highlights the potential of the MERCK SAP formulation as a promising strategy for oxidative stress mitigation in maize under drought conditions, warranting further investigation into its species-specific mechanisms of action.
The efficiency of SAPs is modulated by environmental and soil conditions, which can affect their water retention capacity and overall performance. More et al. [97] reported that SAP absorption capacity is strongly affected by factors such as pH, water type, and soil composition. Furthermore, Abdallah et al. [98] suggested that small-particle SAPs may limit plant access to stored water, which may explain the limited effects observed in this study.
In addition, SAP performance is influenced by cation interactions in the soil. Prior research has demonstrated that different ions affect SAP absorption efficiency, following the trend: K2+ > Na2+ > NH4+ > Al3+ > Fe3+ > Mg2+ > Ca2+ [23,24]. Kaur et al. [11] found that dissolved mineral compounds reduce SAP water retention capacity due to interactions with soil ions, leading to increased Na+ absorption by roots, which can inhibit plant growth [23]. Furthermore, Situ et al. [26] demonstrated that cation exchange between potassium- and sodium-based SAPs can result in excessive accumulation of Na+ and K+, reducing root biomass, length, and area, while also causing deficiencies in Ca2+ and Mg2+, further impairing plant development. The results of the absorption capacity 1 investigation align with these findings, showing that there is an interaction between the applied SAPs and the water mineral content, as well as the cations and anions in the fertilizer. This interaction affects the efficiency of these materials under drought stress conditions.
This study provides a comprehensive evaluation of the effects of SAP application on morphological, physiological, and biochemical traits in rapeseed and maize under controlled drought stress. Despite prior reports suggesting agronomic benefits, our results demonstrate that SAPs confer limited mitigation of drought- induced stress responses, with substantial variability across species and genotypes. SAP treatment did not consistently enhance biomass production or antioxidant capacity. The pronounced genotype-specific variation underscores the necessity of incorporating genetic background as a critical factor when assessing SAP efficacy. Broad, non-specific use of SAPs is unlikely to yield predictable benefits across diverse environmental conditions. The limited effect on antioxidative responses further highlights the inadequacy of current SAP formulations in influencing key drought-adaptive physiological pathways. The interactions between SAPs, soil properties, and plant physiological processes re- main insufficiently understood. Future investigations should integrate detailed soil chemistry, polymer material science, and high-resolution plant phenotyping to characterize these relationships better. Alternative application strategies, such as seed coating, should be explored to improve SAP effectiveness while minimizing unintended physiological costs. Aligning SAP technologies with genotype-specific breeding programs and precision water management may offer more effective solutions. This study establishes a data-driven framework for identifying phenotypic markers associated with SAP responsiveness by applying machine learning approaches for trait prioritization. However, the findings also emphasize that substantial innovation in material composition, application strategies, and genotype-specific optimization is essential before SAPs can be reliably integrated into sustainable drought-resilience efforts. An additional aspect not discussed in this analysis but worthy of investigation in future research is the degradability of the material and its potential toxicological implications upon soil application, especially concerning its interaction with soil microbiota. Moreover, translating these insights from controlled environments to field conditions remains critical for future application.
The following supplementary materials are available online, Figure S1: Shoot height analysis, Figure S2: Root length analysis, Figure S3: Leaf area analysis, Figure S4: Root area analysis, Figure S5: Plant dry weight analysis, Figure S6: Plant fresh weight analysis, Figure S7: Root fresh weight analysis, Figure S8: Root dry weight analysis, Figure S9: Shoot dry weight analysis, Figure S10: Root water content analysis, Figure S11: Shoot water content analysis, which correspond to the phenotypes described in the “Phenotype Analysis” section.
The dataset of the study is available from the authors upon reasonable request.
MG and AOS conceived the study and supervised the research. AA conducted the experiments, performed computational analyses, prepared the datasets, and carried out the literature review. MS and PD contributed to the experimental work and, together with HB and JH, assisted in data interpretation. HB also contributed to data analysis. AA and MG drafted the manuscript. MG led the project management and coordination. All authors reviewed and approved the final version of the manuscript.
The authors declarethat they have no conflicts of interest.
Not applicable.
The publication of this article was funded by the Open Access Publication Fund of the South Westphalia University of Applied Sciences. We would like to thank Martin Wutke, Imran Khan, Muhammad Jawad, Iris Henneken, Katja Johänning, Elena Meininghaus, Albrecht Mühlenschulte, Johanna-Sophie Schlüter, and Cornelia Windhorn for their contributions to this work.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
97.
98.
Abdolmaleki A, Dapprich P, Bertram H, Hollensteiner J, Schmitz M, Schmitt AO, et al. Impact of super absorbent polymers (SAPs) on the morphological, physiological, and biochemical responses of rapeseed and maize under drought stress. Crop Breed Genet Genom. 2025;7(3):e250009. https://doi.org/10.20900/cbgg20250009.

Copyright © Hapres Co., Ltd. Privacy Policy | Terms and Conditions




